Agreement on the International Dolphin Conservation Program (AIDCP)
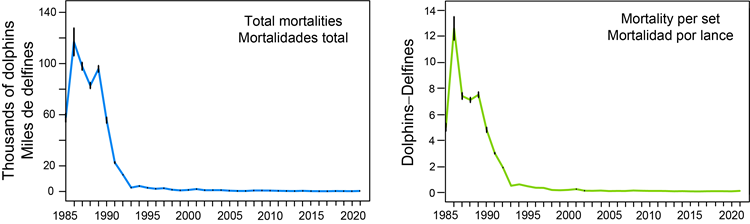
The first objective of the Agreement on the International Dolphin Conservation Program (AIDCP) is to progressively reduce incidental dolphin mortalities in the tuna purse-seine fishery in the Agreement Area to levels approaching zero, through the setting of annual limits.
Topics